
A Scrappy Microbiologist Won't Back Down & Advances His Case
When a man knows something's true, when it's obvious to him, but his peers and his colleagues believe something else, what does he do? Especially if it has to do with a subject considered to be very important, even foundational for the work he does, what does he do?
Does he decide to set his ideas on paper for the whole world to see? This guarantees fierce opposition and sometimes personal attack. To choose this path requires willingness to face that opposition and stand firm. Such intellectual courage is rare but not vanishingly so. This is the story of a man who has that courage.
The Concept of Irreducible Complexity
By the early 1990s, Dr. Michael J. Behe, professor of biochemistry at Lehigh University, had come to doubt the efficacy of Darwinian evolution. (Behe uses the term Darwinian evolution to distinguish it from evolution meaning simply change over time, which is not controversial and which he accepts. Darwinian evolution, on the other hand, is claimed to be the result of unguided, naturalistic processes of random mutation and natural selection, which he sees as severely limited).
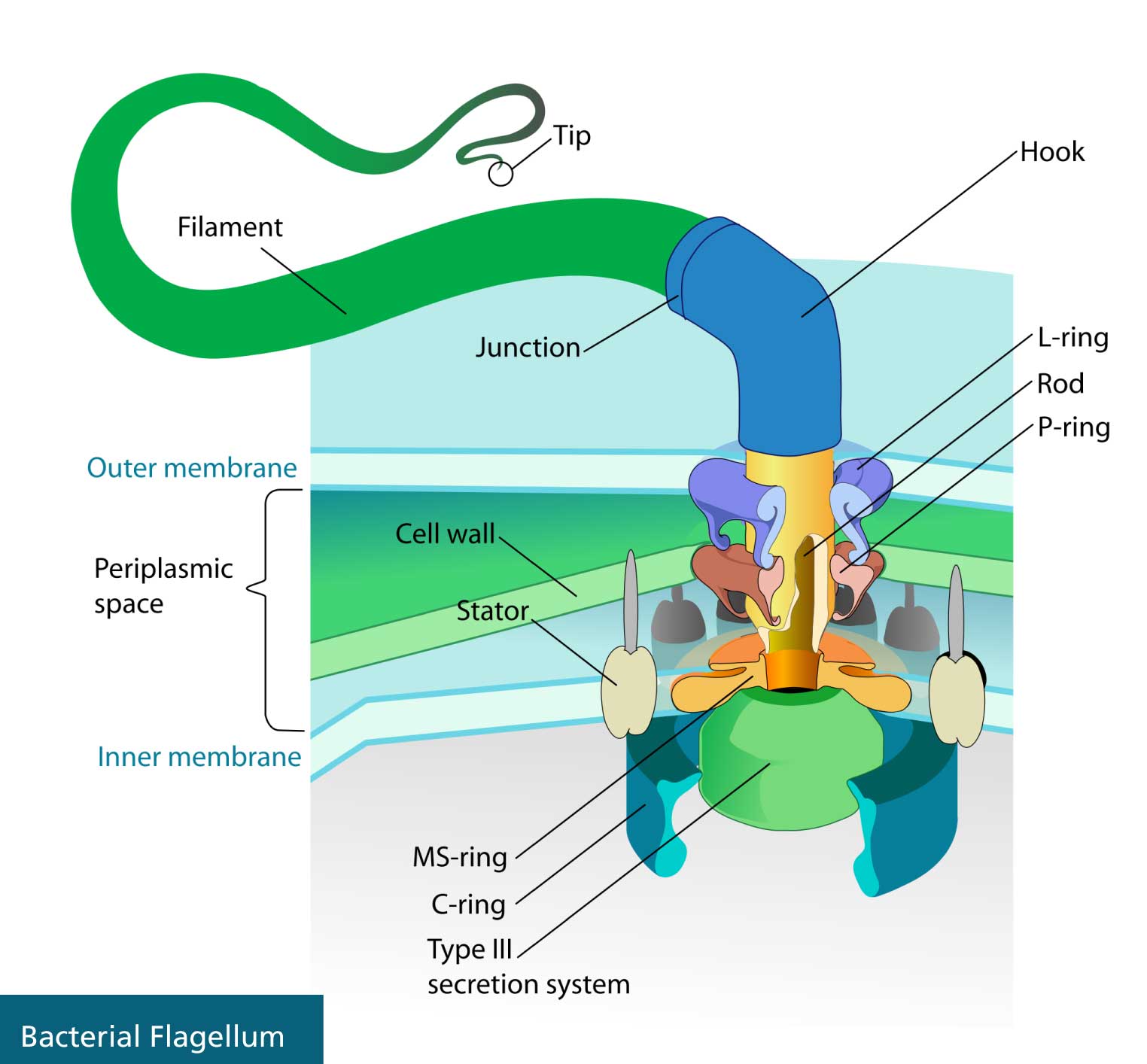
In 1993, Behe attended a meeting with other intelligent design advocates at Pajaro Dunes, California. He presented a now key idea for intelligent design called irreducible complexity. By 1996, he had completed his seminal book, Darwin's Black Box: The Biochemical Challenge to Evolution, in which he introduced irreducible complexity in print. Behe defined it as "a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning." He gave several now-famous examples: the bacterial flagellum, the blood-clotting system, and the immune system.
Behe is a biochemist. His knowledge of the intricacies of biochemical systems led him to his ideas about intelligent design. If there are systems in the cell, or in the body, that are composed of multiple interacting parts, all of which must be present for the system to work, such that there is no function until they are all there, the question arises, How did that system come to be? It is irreducibly complex. It can't evolve one piece at a time and then assemble all at once—hey presto!—with everything in its proper place. It can't build gradually without guidance because it has no function yet. Evolution has no foresight. It doesn't collect parts and store them away for some future use. A non-functional protein will be subject to recurring mutation while it "waits" for other parts to evolve.
The only way for an irreducible, multi-part, well-fitted system to evolve is for it to have multi-functional proteins that can assemble themselves into a new structure, such that the assembly can adapt to a new function gradually. How likely is that? Let's look at the primary test case for Behe's irreducible complexity, the bacterial flagellum.
The Problem with Homology
The flagellum is the means by which the bacterium swims. It consists of a whip-like tail attached to a motor and rod that spin the tail, acting very much like an outboard motor and propeller on a boat. The motor resembles a man-made engine, only it is powered by proton flow rather than gas, with a drive shaft, bushings, stator, and clutch. The parts fit together precisely, and the core parts are essential for function—if one is removed, the whole thing ceases to work. Present-day flagella are composed of 30–50 proteins, depending on the bacterial species. Twenty proteins are part of the core function.
Assembling twenty proteins would take a lot of evolutionary time, one would think. What do Behe's critics say?
Ian Musgrave wrote a piece called"Evolution of the Bacterial Flagellum," published in 2004.1 In it, he claimed there was a path by which the bacterial flagellum might have evolved. The key rhetorical strategy he uses is to identify a sequence of extant organisms that might illustrate an evolutionary pathway, and then declare it to be so. He also uses similarity of function or structure to imply relatedness.
Many evolutionary arguments hinge on that one word, "similarity," or its evolutionary sister, "homology." Homology means having similarity of sequence, structure, or function because of shared ancestry, thereby assuming the thing it is meant to prove.
Some background: There are two different kinds of bacteria: eubacteria and archaebacteria, with flagella that superficially look alike. Both kinds of flagella provide motility but are also involved in secretion. (They have to be, in order to build themselves outside the cell wall.) Notably, these two kinds of flagella are built from different proteins. As Musgrave says, "There is no homology between the flagellar proteins of the eubacteria and archaebacteria." That means there is no shared ancestry between the two kinds of flagella.
To make his case, Musgraveoutlines an evolutionary path for archaebacteria. He takes secretion from some bacteria, slides over to twitching motility in other bacteria, which involves some secretion, and spins up rotary motility in yet other bacteria. He doesn't state how this happened (there are lots of missing steps) or demonstrate how it works; he just asserts it. (This is a not uncommon mode of argumentation in evolutionary biology. If a sequence of organisms can be found that might illustrate a path to some structure or function, then that perhaps-possible-pathway is taken as a demonstration that evolution of that structure or function is inevitable.) He asserts without evidence that
The archaebacterial flagellum passed from being a secretory structure, to a gliding motility system, to a rotatory swimming system. At each point there was time for substructures to adapt to each other before the next stage.
That's artful storytelling, rather than fact.
Turning to the eubacterial flagellum, Musgrave says first that the majority of proteins in this flagellum have homology with other proteins outside the flagellum.
[We] now know that the majority of the eubacterial flagellar proteins have homologs. As I have pointed out, the motor proteins and the chemical-sensing system have homologs in other secretory systems. Other flagellar proteins such as the sigma factors have homologs as well. In the end there is not much unique left in the flagellum.
Here he's sneaking in an evolutionary story through the use of the word "homology." All it really means is that there is some similarity in sequence with non-flagellar proteins. But just because two proteins look alike does not mean there is a path between them.
Then comes the sleight of hand. Musgrave draws a parallel between archaebacterial and eubacterial flagellar evolution, based on archaebacteria's putative evolutionary sequence as described above, even though they are unrelated. First, a secretory system arose in eubacteria. Next, an ion pump (which later became the motor protein) improved secretion. Then the primitive flagellar filament arose as part of the protein secretion structure. Gliding/twitching motility came next (arose) and was refined into swimming motility. In his text that word "arose" is used as if these proteins rose up out of the dawn mists fully formed, like Aphrodite.
Memorize this: Just because two proteins look alike does not mean there is a path between them. Just because creatures look like they could be related in a functional series, does not mean they are so related. Just because an evolutionary biologist says he has a plausible pathway doesn't make it so, either.
Evolution by Loss of Function
I will pass over other controversies involving the other irreducibly complex systems that Behe describes in Darwin's Black Box, and move on. Behe's next book, The Edge of Evolution: The Search for the Limits of Darwinism, was published in 2008. In it, he explores the subject of his postdoctoral work, malaria.
Malaria is a dreadful disease, and the parasite that causes it is very good at evading anti-malarial drugs. The drug chloroquine, however, had a longer run of effectiveness than any other. Behe said that because of the number of malarial parasites in the human population, if all it took for a parasite to develop resistance to a certain medication were one mutation, such resistance would develop quickly. But if two mutations were required, it would take decades. And that is how long it took for chloroquine resistance to develop. Behe predicted that when the molecular basis for the resistance was discovered, it would be shown that two mutations were responsible.
He was right. A paper published in the Proceedings of the National Academy of Sciences, which he quotes, says,
A minimum of two mutations sufficed for (low) CQ transport activity, and as few as four conferred full activity. . . . The findings presented here reveal that the minimum requirement for (low) CQ transport activity in both the ET and TD lineages of CQR PfCRT is two mutations.2
In 2010, Behe published a paper in the Quarterly Review of Biology called "Experimental Evolution, Loss-of-Function Mutations, and the First Rule of Adaptive Evolution,"3 in which he summarized years of work by geneticists on numerous bacteria and viruses, and laid out this principle: "Break or blunt any functional coded element whose loss would yield a net fitness gain." In other words, if an evolutionary problem can be solved by reducing or eliminating some genetic function, that is the path most likely to be taken.
Much of evolution occurs by loss of function, as long as the breaking or blunting of a coded functional element (which results in loss of function) actually helps the organism survive or reproduce better. The reason is simple. It is much easier to damage or destroy an existing function than it is to build a new one. But this is a one-way road. Many kinds of mutation are irreversible.
In the years since that paper was published, and with the acceleration of genome sequencing, more and more evidence of evolution by loss-of-function mutation has accumulated.4 Behe was right again.
New Book, New Challenge
Behe is not in a defensive posture, despite his critics. He continues to advance. He has taken his ideas (irreducible complexity, the limits of evolution, and loss of function as the main mode of evolution) to the next level in his new book, just released in February, called Darwin Devolves.5 There he asks, What exactly can Darwinian evolution accomplish, when loss of function is the main road to effecting change, when there is a limit to how far random mutation can go, and when the requirements of irreducible complexity have to be fulfilled?
I will not disclose the answer, but I can guarantee his answer will challenge—and no doubt aggravate—his critics.
Notes
1. In Why Intelligent Design Fails: A Scientific Critique of the New Creationism, Taner Edis and Matt Young, eds. (Rutgers Univ. Press, 2004).
2. Ann Gauger, "A Pretty Sharp Edge," Evolution News (July 28, 2014): https://evolutionnews.org/2014/07/a_pretty_sharp; "Best of Behe," Evolution News (Nov. 10, 2016): https://evolutionnews.org/2016/11/best_of_behe_a; Summers et al., "Diverse mutational pathways converge on saturable chloroquine transport via the malaria parasite's chloroquine resistance transporter," PNAS, (Apr. 29, 2014): pnas.org/content/111/17/E1759.
3. Michael J. Behe, "Experimental evolution, loss-of-function mutations, and 'the first rule of adaptive evolution,'" The Quarterly Review of Biology (December 2010): journals.uchicago.edu/doi/abs/10.1086/656902?journalCode=qrb.
4. Ann Gauger, "Bacteriophages, Budding Yeast, and Behe's Vindication," Evolution News (Nov. 26, 2018): https://evolutionnews.org/2018/11/bacteriophages-budding-yeast-and-behes-vindication.
5. Michael J. Behe, Darwin Devolves: The New Science About DNA That Challenges Evolution (HarperCollins, 2019).
is the Director of Science Communication at the Center for Science and Culture at the Discovery Institute and a senior research scientist at Biologic Institute.
Get Salvo in your inbox! This article originally appeared in Salvo, Issue #48, Spring 2019 Copyright © 2026 Salvo | www.salvomag.com https://salvomag.com/article/salvo48/behe-right-or-wrong

















